强化生物除磷技术(enhanced biological phosphorus removal,ebpr)由于经济、高效的特点被广泛应用于各大城市污水处理厂.通过分子生物技术已经确认菌群β-proteobacteria是ebpr活性污泥中的优势聚磷菌(paos),将其命名为c and idatus accumulibacter(hesselmann et al., 1999;crocetti et al., 2000).accumulibacter 是目前污水生物处理领域广泛认可并且研究最多的一种paos(zilles et al., 2002).ppk1基因可以编码合成poly-p的聚合磷酸盐激酶(ppk1),其进化速度是16s rrna 的5倍,是研究accumulibacter各进化分支很好的基因标记物.mcmahon等(2007)基于ppk1的系统发育分析将type i划分为type i和type ii,其中,type i 进一步划分为ia、ib、ic、id、ie 5个分支;type ii划分为iia、iib、iic、iid、iie、iif、iig 7个分支.
现有研究证实,accumulibacter在污水厂存在很高的多样性,且主要分布于typeⅱ,但水厂运行与各分支分布的相关性不清楚.宏基因组的研究也证实了分支多样性,这种多样性与污水处理厂的稳定性有一定关系(albertsen et al., 2012).mielczarek等(2013)对丹麦28个污水处理厂的研究发现,ia和iic分支几乎出现在每个处理厂.peterson等(2008)对美国几个污水处理厂及一些淡水湖泊、河口沉积物样品进行研究发现,iid存在于大部分的淡水、河口沉积物中及少部分污水处理厂中,有的湖泊淡水中仅发现iid 1个分支.由此可见,来自不同地域的污水处理厂,accumulibacter进化枝的分布有明显差异.对实际处理系统中accumulibacter的研究主要发现了ia、iia、iib、iic、iid、iif这6个分支,尤其是前5个分支的研究报道较多(he et al., 2007; gebremariam et al., 2011; mcmahon et al., 2007; mielczarek et al., 2013),iif较少存在.而在本研究中iif在几个污水处理系统中含量丰富,首次作为重点研究.其他6个分支(ib、ic、id、ie、iie、iig)主要存在于自然界的淡水、湖泊、河流沉积物等地方,很少在实际污水处理厂中出现(mcmahon et al., 2007).因此,着重关注ia、iia、iib、iic、iid、iif这6个分支而不是整体的12个分支.关于中国城市污水处理厂中accumulibacter 6个分支的菌群结构及定量分析的研究尚未见报道.
近年来,许多研究者针对污水处理系统的运行参数和accumulibacter各分支的组成分布的关系展开研究.关于电子受体的影响,kim等(2013)认为所有杆状的accumulibacter分支,包括分支iic、 ia和iif在具有足够的硝酸盐还原活性的污泥中可以成功的利用硝酸盐吸磷.flowers等(2009)发现反硝化吸磷时,ia、iia分别以硝酸盐、亚硝酸盐作电子受体.flowers等(2013)在最近的研究中发现,在动力学上clade iia活性与温度正相关,而ia与温度呈负相关.迄今为止,究竟有哪些因素主导了实际污水处理系统accumulibacter的种群结构和动态变化仍然没有确切的研究结果.
本研究选择9个具有代表性的大型市政污水处理厂,分析不同运行状况的污水处理厂活性污泥中c and idatus accumulibacter主要分支(i、iia、iib、iic、iid、iif)的丰度及分布特点,揭示污水处理厂c and idatus accumulibacter进化枝水平菌群结构、丰度与工艺运行的相关性.
2 材料与方法
2.1 城市污水处理厂的活性污泥样品
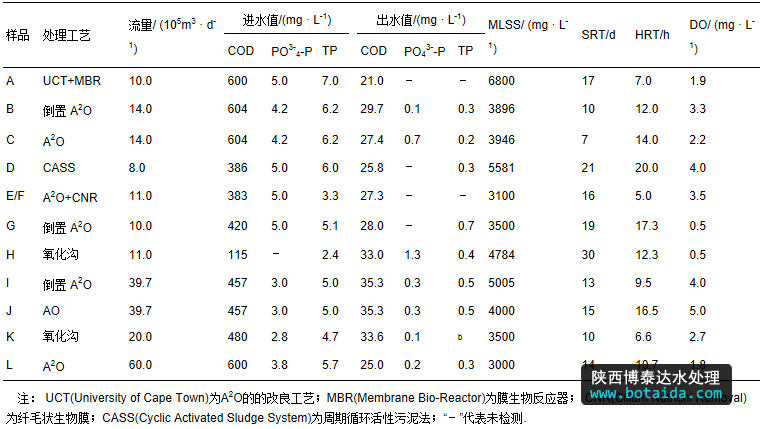
实验所用的污泥样品分别采集于9个污水处理厂,其中有2个水厂各包含2个工艺,共11个工艺.每个工艺取1个污泥样品.其中,针对a2o 纤毛状生物膜(cnr)工艺取2个样品,分别位于加膜区和无膜区,是为了考察加膜对聚磷菌的菌群结构是否有影响,因此,共有12个样品.12个样品依次命名为a、b、c、d、e、f、g、h、i、j、k和l.其中,b和c取自同一水厂,i和j取自同一水厂,e和f取自a2o 纤毛状生物膜工艺的加膜区和无膜区.污水处理厂的运行参数及进出水指标见表 1.
表1 活性污泥样品对应的污水处理系统的运行参数及进出水指
2.2 dna提取、pcr、克隆测序
用1×pbs(0.01 mol · l-1,ph=7.2~7.4)清洗污泥样品3次,14000×g离心2 min,去除上清液,置于-20 ℃保存.采用试剂盒(fast dna spin kit for soil,mp,usa)对dna进行提取.提取后的dna 通过nanodrop spectrophotometer nd-1000(thermo fisher scientic,usa)测量核酸浓度及纯度.
特异性正向引物ppk1-254f(tcaccaccgacggcaagac)和反向引物ppk1-1376r(acgatcatcagcatcttggc)用来扩增总c and idatus accumulibacter ppk1 基因,片段长度为1123 bp.pcr反应采用试剂盒(promega gotaq green master mix,usa),反应体系为25 μl:12.5 μl gotaq green master mix,1 μl(10 mmol · l-1)正向引物,1 μl(10 mmol · l-1)反向引物,0.5~2 μl dna模板,剩余体系用纯水补齐.pcr程序包括:95 ℃预变性10 min;95 ℃变性45 s,61 ℃退火1 min,72 ℃延伸2 min,35个循环;最后在72 ℃反应5 min.
获得的pcr产物经琼脂糖凝胶电泳(agarose ms-6,takara,japan)检测,为单一的目的条带,切胶,用takara agarose gel dna purication kit ver. 2.0(takara,dalian,china)纯化.得到的dna用试剂盒(zero background ta topoisomerase cloning kit,clonesmarter,usa)进行连接转化.连接体系为10 μl,包括1 μl pcloneez-topo载体,1 μl 10×enhancer,0.5~8.0 μl dna,一定量的纯水补齐体系.连接反应完成后将产物加入到感受态细胞dh5a(中美泰和,国产)中进行转化.每个样品随机挑出一定量的阳性克隆子进行测序,构建克隆文库.
2.3 克隆文库建立和系统发育分析
构建文库的序列通过mothur 软件按照 97%相似度进行 otu 划分,将每个 otu 的代表序列与ncbi 数据库中下载的相似性最高最具代表性的菌株序列一起进行比对.采用 mega5.0 利用邻接法(neighbor joining method)进行系统发育分析,通过自举分析方法(bootstrap)检验系统发育树各分支置信度,重复1000次.
2.4 实时定量pcr(qpcr)
采用特异性引物对c and idatus accumulibacter主要的6个分支(i、iia、iib、iic、iid、iif)的ppk1片段和全菌的16s rrna进行qpcr扩增(he et al., 2007; ong et al., 2014).反应在mx3005p实时定量pcr扩增仪(agilent technologies,american)上进行,采用takara sybr premix ex taq kit进行反应,体系为25 μl包括12.5 μl的sybr缓冲液,正反向引物各1 μl(10 mmol · l-1),0.5 μl rox,dna模板2 μl,剩余体系用纯水补齐.特异性引物序列及扩增程序见表 2.
表2 qpcr各分支的特异性引物序列及扩增程序
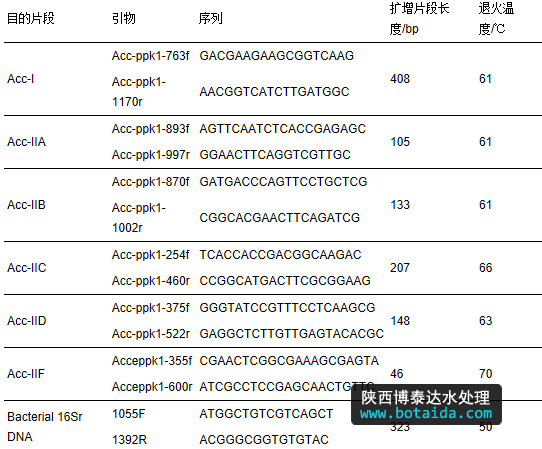
本研究采用质粒法制作标准曲线.c and idatus accumulibacter 6个分支ia、iia、iib、iic、iid、iif实时荧光定量 pcr 标准曲线中的初始模板浓度与ct值之间呈现较好的线性关系,r2在0.990~1.000之间,扩增效率在92%~109.7%之间.
2.5 登录号
测得的序列上传至 genbank 数据库,获得的聚磷菌的序列登录号为:kp147989~kp148181、kt024045~ kt024419.
3 结果与讨论
3.1 12个活性污泥样品中c and idatus accumulibacter 的定量分析
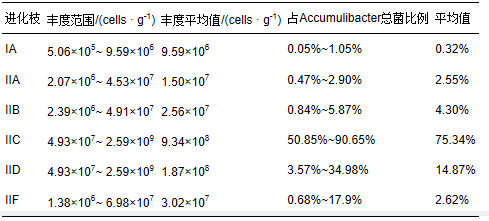
采用实时定量pcr定量分析accumulibacter(i、iia、iib、iic、iid、iif)6个分支的丰度,结果如图 1所示.accumulibacter 6个分支均为单拷贝基因,即accumulibacter的细胞数为ppk1基因的拷贝数.总accumulibacter的定量变化范围为7.32×107(d)~3.05×109 cells · g-1(l),平均为1.20×109 cells · g-1(以mlss计).在12个样品中,只有d样品数量级为107,比其他样品低1~2个数量级,其中的accumulibacter ppk1基因含量偏低;有6个样品数量级为108,分别为h、b、c、i、j和k;样品a、e、g、f和l的数量级为109,这5个样品中的accumulibacter的ppk1基因含量高于其他样品,其中样品l(a2o工艺)含量最高,达到了3.05×109 cells · g-1(以mlss计).12个样品中accumulibacter占总菌的百分比变化范围为0.58%~ 22.77%,平均为9.17%(图 1).
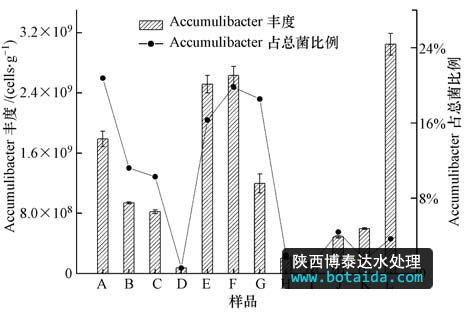
图1 accumulibacter丰度及占总菌的比例
mielczarek等(2013)通过fish技术对丹麦28个城市污水处理厂中accumulibacter进行研究时发现,accumulibacter占总菌的平均比例为3.7%,低于本研究结果.而he等(2008)对5个污水处理厂中accumulibacter占总菌比例的研究结果为9%~24%,与本研究结果基本一致.
从图 1中可以看出,样品a、e、f、g和l中accumulibacter的丰度均在109 cells · g-1(以mlss计)以上,而这5个样品中,l为a2o工艺、a为a2o mbr工艺、e、f为a2o nar工艺,样品g为倒置a2o工艺,且在5个样品中丰度偏低.由此可见,a2o工艺及其改良工艺中accumulibacter的丰度最高.且来自同一工艺的样品e和f证实了在a2o工艺的基础上加膜不会对accumulibacter的丰度造成影响.同为氧化沟工艺的样品h和样品k中accumulibacter的丰度在同一数量级上,为108 cells · g-1(以mlss计).ao工艺的样品j中accumulibacter的丰度和样品h(氧化沟)在同一数量级,和样品g同为倒置a2o工艺的样品i要比样品g小一个数量级.由此判断,倒置a2o工艺和氧化沟工艺的accumulibacter丰度也处于比较高的水平,只有cass工艺对应的样品中accumulibacter丰度明显低于其他工艺.各个样品中accumulibacter占总菌的比例与各个样品accumulibacter的丰度变化趋势基本保持一致.
accumulibacter 6个分支定量结果见图 2.6个分支在12个污泥样品中丰度变化范围及各分支占总accumulibacter的变化范围见表 3.从图 2可以看出,accumulibacter的6个分支中,iic和iid处于同一数量级,为108 cells · g-1(以mlss计),分别占accumulibacter总菌的比例为75.34%和14.87%,是污水处理系统中的优势菌属.尤其是iic丰度达到了109 cells · g-1(以mlss计). iia、iib和iif处于同一数量级,为107cells · g-1(以mlss计),比iic和iid低一个数量级,这3个分支之和占accumulibacter 总菌的比例为9.34%.ia含量最少,数量级比iic和iid低2个数量级,占accumulibacter总菌的比例仅为0.32%.accumulibacter的 6个分支均存在于每个工艺中,但type i所占比例不到1%,而type ii接近100%.各个分支的变化基本和总accumulibacter变化趋势一致,与mielczarek等(2013)的研究结论一致,不同的是mielczarek等没有发现任何处理厂含有accumulibacter的5个分支(ia~iid). 本研究首次报道了iic在9个城市污 水处理厂中丰度最高,ia在9个水厂中丰度最低,accumulibacter的这种群落分布特点明显不同于其他污水处理厂.type ii中的6个分支在12个污泥样品中呈现不同的分布特点,定量结果证实了城市污水处理厂accumulibacter进化枝水平分布的多样性.
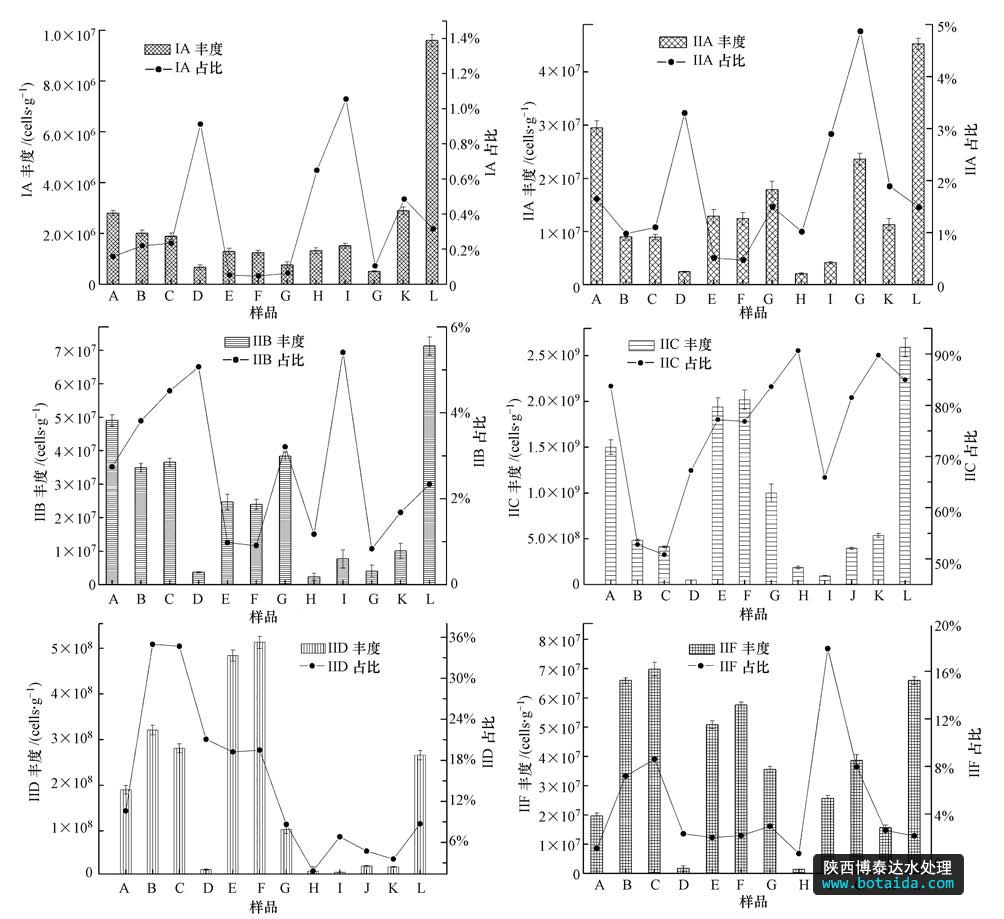
图2 accumulibacter各进化分支丰度及相对含量
表3 12个样品中c and idatus accumulibacter中各进化分支的比例
3.2 12个活性污泥样品中c and idatus accumulibacter 基于otu的多样性分析
9个污水处理厂,共12个样品,构建12个ppk1基因克隆文库进行分析.12个克隆文库的good 覆盖率、chao1 丰富度估计、otu 值与估计的 chao1 值之比(otuobserved/otuestimated)、香农指数见表 4.12个污泥样品的ppk1基因的otus分布如图 3所示.如表 4所示,12个样品的good覆盖率变化范围为71.43%~98.15%.计算的 otu 值与估计的 chao1 值之比除了样品a为47.06%,其余11个样品在62%~100%之间.mehlig等(2013)对国外8个污水处理厂进行了研究,good覆盖率只有35.4%~65.4%.说明本研究的12个克隆文库可以代表 12个样品中accumulibacter的群落组成.样品a可能需要测更多的数据保证数据的可信度.
表 4同时给出了12个 ppk1 基因克隆文库中的香农指数,根据表中结果发现,样品c只有1个otu,说明该样品中ppk1基因多样性很低,香农指数为零.另外11个样品的香农指数在0.15~2.04之间,其中,样品a、d、g、j和k含有较丰富的多样性.he等(2007)对9个市政污水处理厂中accumulibacter的群落进行研究,香农指数在0.2~1.4之间.与本研究对12个克隆文库中accumulibacter的群落多样性研究相符合.
表4 克隆文库中序列多样性及覆盖率
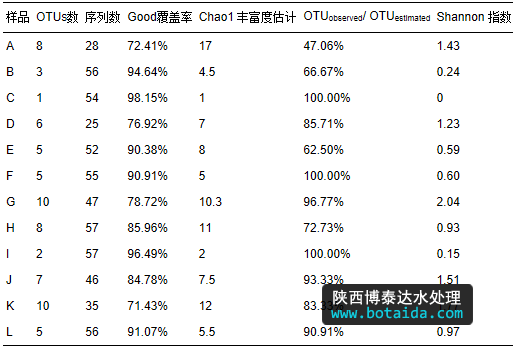
12 个ppk1基因克隆文库中共得到568 条 ppk1序列,按照 3% cut-off的距离划分为 29个 otus. 从图 3可以看出,样品c仅有1个otu,意味着样品c中accumulibacter的多样性最不丰富.而样品b和样品i分别包含3、2个otus,说明这2个样品中accumulibacter的多样性水平比较低.样品a、d、g、j、h和k分别含有8、6、10、8、7和10个otus,具有较高的多样性水平,样品e、f和l均含有5个otus,较前6个样品多样性偏低.
otu1共出现12次,即每个样品中都有序列出现在otu1中,同时还是样品b、c、e、f、f、i和j共6个克隆文库中的优势otus.otu3和otu5在6个样品中出现,尤其在样品h的克隆文库中分布广泛.otu7出现了4次.有4个otus共出现3次,分别为otu2、otu4、otu6和otu11.上述8个otus为accumulibacter的主要种属.有10个otus(otu10、otu12、otu13、otu15、otu17~ otu20、otu22、otu23)出现2次,有11个(otu9、otu10、otu14、otu16、otu21、otu24~ otu29)共出现1次,这21个otus中每个otus包含1~2条ppk1序列,不是accumulibacter的主要种属.
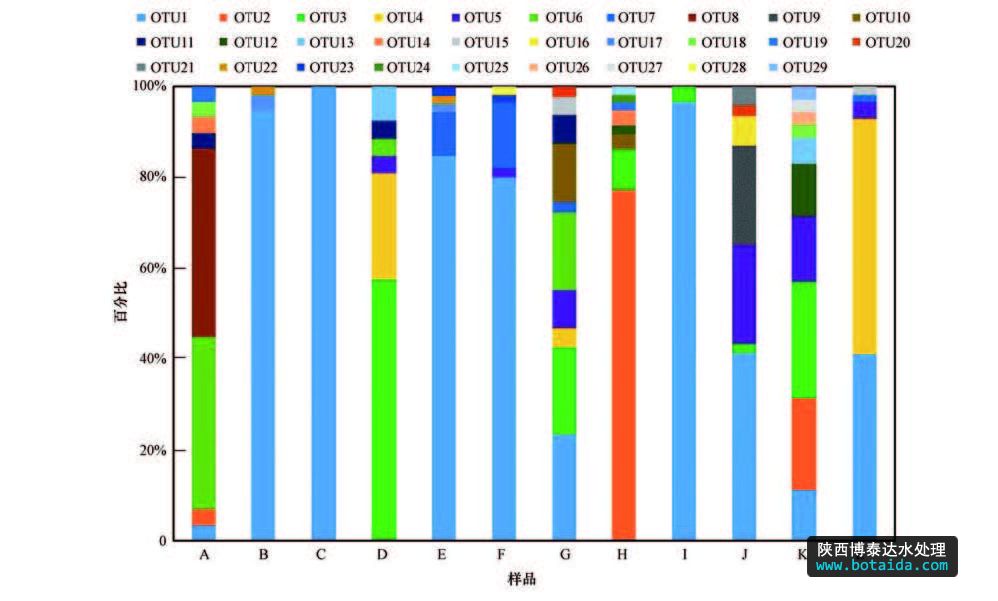
图3 12个污泥样品的ppk基因的otu分布图
3.3 12个活性污泥样品中c and idatus accumulibacter基于系统发育树的多样性分析
利用29个otus的代表序列建立的系统发育树如图 4所示.he等(2007)对9个市政污水处理厂中accumulibacter的群落进行分析认为,实际污水处理厂污水组成和运行情况比较复杂会导致污水中accumulibacter种类比较丰富,所测得的序列至少分布在3个进化枝中.type i包括ia、ib、ic进化枝,type ii包括iia、iib、iic、iid、iif进化枝.id、ie、iie和iig在淡水和河口沉积物中发现,在此不做考虑.另外,序列rhodocyclus tenuis 作为参考序列.以往的研究均是通过各个水厂中的代表序列建系统发育树,但本研究中采用各otus中代表序列建立系统发育树,这样更方便从整体上观察几个工艺中accumulibacter的分布情况.如图 4所示,otus在几个进化枝间分布比较均匀,有27个otus属于type ii型的accumulibacter,仅有2个otus(共12条序列)属于type i型,说明实际污水除磷系统中的accumulibacter主要以type ii型为主,与qpcr结果一致.
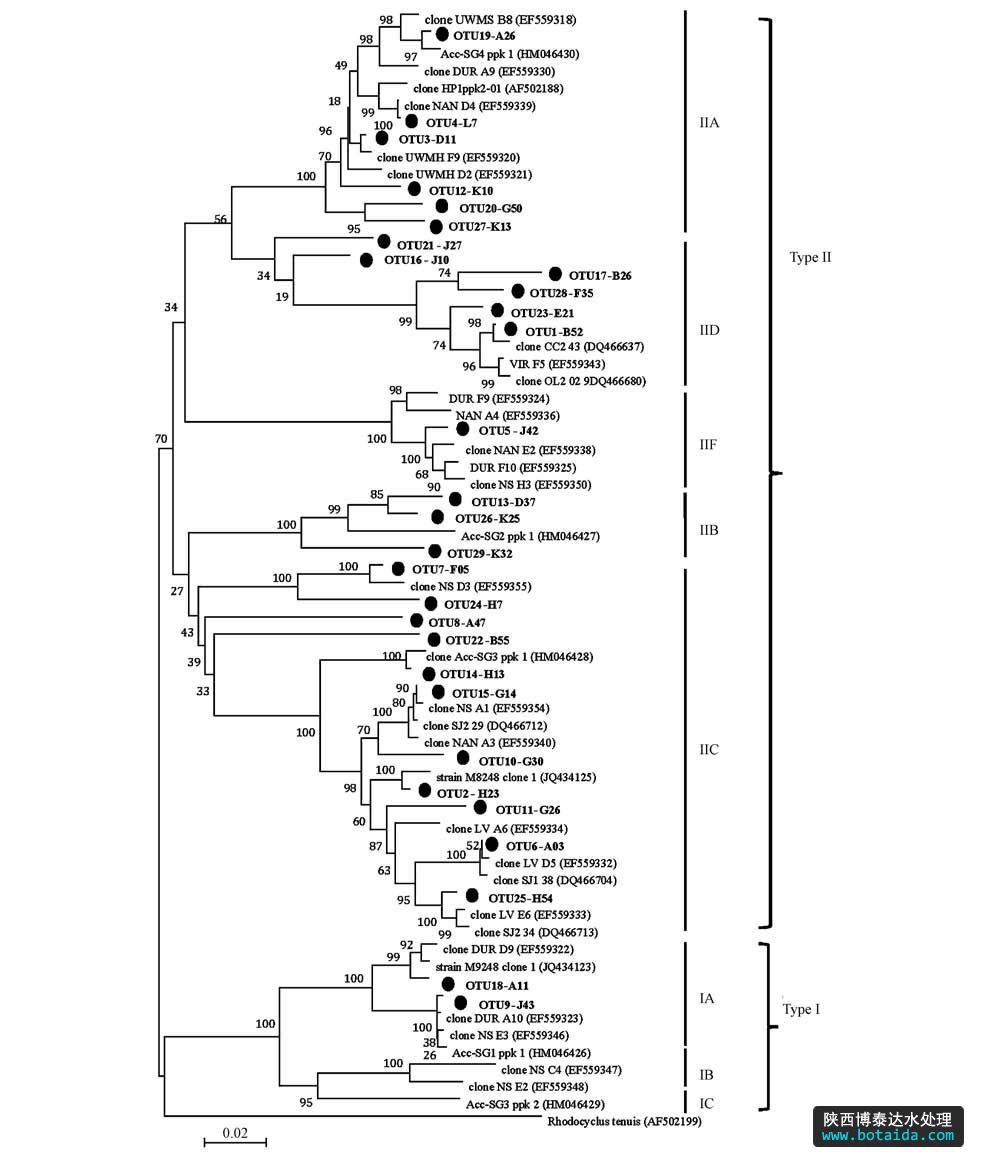
图4 基于ppk1基因的系统发育树
结合图 4和图 5可知,iia包含6个otus(88条序列),iib包含3个otus(6条序列),iic包含11个otus(124条序列),iid包含4个otus(315条序列),iif包含1个otu(共23条序列).其中,优势otus(otu1~otu3)分别隶属于iid、iic、iia.qpcr和pcr-cloning-sequencing的研究结果都证实了12种污水处理工艺中type i丰度很低,主要以 type ii为主,且iia、iic、iid是系统中的优势分支.
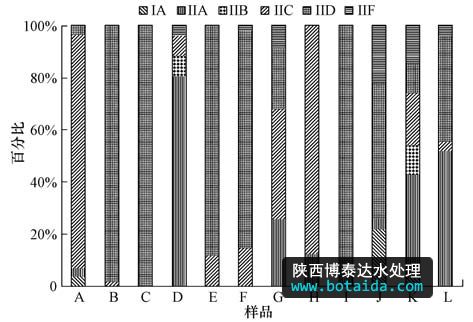
图5 12个污泥样品中accumulibacter 6个分支分布图
3.4 c and idatus accumulibacter各进化枝的分布与ebpr性能之间的关系
结合12个污泥样品的qpcr分析和克隆文库的分析可知,type i和type ii均存在于城市污水处理厂中,以type ii为主,type i占accumulibacter总量的比例低于0.32%.type ii中以iic为主,iid、iia次之,iib含量较不丰富,而ia丰度最小.这与实验室规模的反应器中accumulibacter的种群结构明显不同,实验室反应器富集的accumulibacter主要以ia和iia为主(he et al., 2010;gonzalez-gil et al., 2011).对于污水处理厂中accumulibacter各进化枝的分布与ebpr性能之间的关系分析如下:
1)除磷效果不好可能和iid占总accumulibacter比例较高有关.本课题组前期对实验室规模的muct反应器中accumulibacter种群进行反硝化除磷的研究,发现在以no-2为电子受体的反硝化除磷系统中iid始终是优势菌属,占总accumulibacter的90%以上(曾薇等,2013),说明iid分支适合于以no-2为电子受体进行缺氧吸磷.而实际污水处理厂是以o2为电子受体的好氧吸磷及以no-3为电子受体的缺氧吸磷,iid比例的升高导致除磷效果变差.
2)关于iic在污水处理厂中的功能还需要进一步的研究.有报道称iic可以利用硝酸盐为电子受体进行缺氧吸磷(kim et al., 2013),但也有报道称iic是导致ebpr系统恶化的原因(slater et al., 2010).由此可见,在系统发育上属于同一进化枝的accumulibacter由于工艺和运行条件的不同可能会具有不同的功能.
4 结论
1)qpcr结果表明,accumulibacter 6个进化枝(ia、iia、iib、iic、iid、iif)存在于12个污泥样品中,占污水处理系统中总菌的比例平均为9.17%.a2o工艺及其改良工艺中总accumulibacter丰度最高,占到总菌的22.8%,具有较好的生物除磷性能.分支iic在11种工艺中均丰度最高,平均占总accumulibacter比例的75.34%.iid次之,占总accumulibacter的比例平均为14.87%,ia丰度最低,占总accumulibacter比例平均只有0.32%.
2)12个活性污泥样品的系统发育分析也证实了accumulibacter以type ii 为主,type i少量存在.typeii中以iic、iid、iia为主,iib、iif丰度偏低.
3)污水处理厂中accumulibacter各进化枝的分布与ebpr性能之间存在一定关系.除磷效果不好可能和以亚硝酸盐为电子受体的iid占总accumulibacter比例较高有关.而iic在污水处理厂中的功能还需要进一步的研究.